Cell-cell adhesion
To form an anchor junction, cells must first adhere. Next, a bulky cytoskeletal apparatus must be assembled around the molecules that directly mediate adhesion. The result is a well-defined structure (a desmosome, hemidesmosome, focal adhesion, or adherens junction) that is easily identified by electron microscopy. Indeed, electron microscopy provided the basis for the original classification of cell junctions. However, in the early stages of cell junction development, before the cytoskeletal apparatus has been assembled, cells often adhere to one another without clearly showing these characteristic structures; in the electron microscope, one can simply see two plasma membranes separated by a small gap of defined width.
Functional tests show, however, that the two cells are stuck together, and biochemical analyzes can reveal the molecules responsible for the adhesion. The study of cell-cell junctions and the study of cell-cell adhesion Recombinants were once quite distinct endeavours, originating from two different experimental approaches: junctions through electron microscopic description and adhesion through functional testing. and biochemistry. Only in recent years have these two approaches begun to converge into a unified view of the molecular basis of cell junctions and cell adhesion.
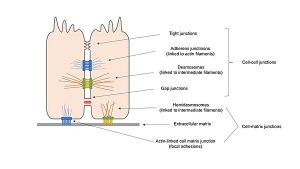
In the previous section, we focused on the structures of mature cell junctions. In this section, we turn to functional and biochemical studies of the cell-cell adhesion mechanisms that operate when cells migrate over other cells and when they assemble into tissues, mechanisms that precede the construction of mature cell-cell anchor junctions. We start with a critical question for embryonic development: what mechanisms ensure that a cell attaches itself to the appropriate neighbours at the right time?
Dissociated vertebrate cells can reassemble into organized tissues through selective cell-cell adhesion
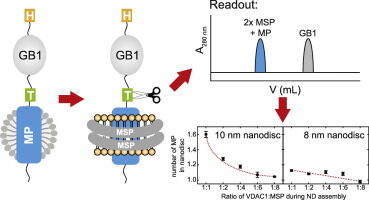
Unlike adult vertebrate tissues, which are difficult to dissociate, embryonic vertebrate tissues dissociate easily. This is usually done by treating the tissue with low concentrations of a proteolytic enzyme such as trypsin, sometimes combined with removal of extracellular Ca2+ and Mg2+ with a divalent cation chelator (such as EDTA). These reagents disrupt the protein-protein interactions (many of which rely on divalent cations) that hold cells together. Surprisingly, dissociated cells often reassemble in vitro into structures that resemble the original tissue.
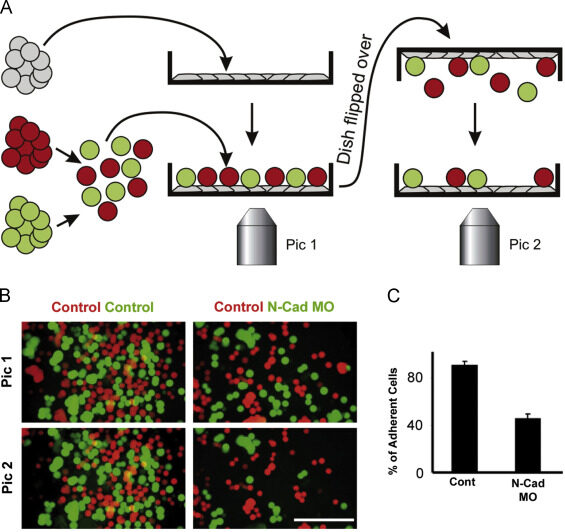
Such findings reveal that tissue structure is not just a product of history; it is actively maintained and stabilized by the system of affinities that cells have with each other and with the extracellular matrix. A striking example of this phenomenon is seen when dissociated cells from two vertebrate embryonic organs, such as the liver and the retina, are artificially mixed and formed into a granule: the mixed aggregates gradually separate according to their organ of origin. More generally, disaggregated cells are found to adhere more readily to aggregates from their own organ than to aggregates from other organs.
Obviously, there are cell-cell recognition systems that make cells of the same differentiated tissue preferentially adhere to each other; these adhesive preferences are presumably important in stabilizing tissue architecture. Cells adhere to each other and to the extracellular matrix through cell surface proteins called cell adhesion molecules (CAMs), a category that includes the transmembrane adhesion proteins discussed earlier. The CAMs can be cell-cell adhesion molecules or cell-matrix adhesion molecules. Some CAMs are Ca2+ dependent, while others are Ca2+ independent.
Ca2+-dependent CAMs appear to be primarily responsible for the tissue-specific cell-cell adhesion seen in early vertebrate embryos, which explains why these cells can be disrupted by Ca2+-chelating agents. CAMs were initially identified by creating antibodies against cell surface molecules and then testing the ability of the antibodies to inhibit cell-cell adhesion in a test tube. These rare adhesion-inhibiting antibodies were then used to characterize and isolate the adhesion molecule recognized by the antibodies.
Non-junctional contacts can initiate cell-cell adhesions which junctional contacts then orient and stabilize
We have seen that adhesive contacts between cells play a crucial role in organizing tissue and organ formation in developing embryos or in adult tissues that are repaired after injury. Very often, these contacts do not involve the formation of organized intercellular junctions that appear as specialized structures in the electron microscope. The interacting plasma membranes are simply seen to approach and run parallel, separated by a gap of 10 to 20 nm. This type of “non-attachment” contact may be optimal for cell locomotion: close enough to provide traction and allow transmembrane adhesion proteins to interact, but not so tight or anchored so solidly to the cytoskeleton as to immobilize the cell.
A reasonable hypothesis is that unbound cell-cell adhesion proteins initiate cell-cell adhesions, which are then oriented and stabilized by the assembly of complete intercellular junctions. Many of the transmembrane proteins involved can diffuse in the plane of the plasma membrane and, in this or other ways, can be recruited to sites of cell-cell (and cell-matrix) contact, allowing adhesions that do not adhere. unite enlarge and mature into union adhesions. . This has been shown with some integrins and cadherins, which help initiate cell adhesion and then become integral parts of cell junctions.
The migratory tip of an axon, for example, has a uniform distribution of cadherins on its surface, which helps it stick to other cells along the migration path. It also has an intracellular reservoir of cadherins in vesicles just below the plasma membrane. When the axon reaches its target cell, it is thought to release intracellular cadherin molecules at the cell surface, where they help form a stable contact, which matures into a chemical synapse. As discussed above, antibodies against adherens junction proteins block the formation of tight junctions as well as adherens junctions, suggesting that the assembly of one type of junction may be a prerequisite for the formation of another.
An increasing number of monoclonal antibodies and peptide fragments have been produced that can block a single type of cell adhesion molecule. In addition, an increasing number of genes encoding these cell surface proteins have been identified, creating new opportunities to manipulate the adhesive machinery of cells in culture and in experimental animals. It is now possible, therefore, to inactivate the various cell-cell adhesion proteins in combinations, a requirement for deciphering the rules of cell-cell attachment and recognition used to build complex tissues.
Summary
Dissociated cells from various tissues of vertebrate embryos preferentially reassociate with cells from the same tissue when mixed. This tissue-specific recognition process in vertebrates is primarily mediated by a family of Ca2+-dependent cell-cell adhesion proteins called cadherins, which hold cells together through homophilic interaction between these transmembrane proteins on adjacent cells. For this interaction to be effective, the cytoplasmic part of the cadherins must be linked to the cytoskeleton by cytoplasmic anchoring proteins called catenins.
Two other families of transmembrane adhesion proteins have important roles in cell-cell adhesion. Selectins function in transient Ca2+-dependent cell-cell adhesions in the bloodstream by binding to specific oligosaccharides on the surface of another cell. Members of the immunoglobulin superfamily, including N-CAM, mediate Ca2+-independent cell-cell adhesion processes that are especially important during neuronal development.
Even a single cell type uses multiple molecular mechanisms to adhere to other cells (and to the extracellular matrix). Therefore, the specificity of cell-cell (and cell-matrix) adhesion observed in embryonic development must result from the integration of several different adhesion systems, some of which are associated with specialized cell junctions, while others are not.